Head Direction Cells: The Brain's Internal Compass
In 1983, James Ranck Jr. found neurons in the rat dorsal presubiculum that fire whenever the animal's head points in one specific compass direction — regardless of where in the room the animal stands. This article explains how head direction cells work, what their tuning curves reveal, how the vestibular-to-cortex circuit generates and sustains the directional signal, and why the ring attractor model built to explain them became a cornerstone of computational neuroscience.

June 15, 2026 · 8:26 AM
1 subscriptions · 19 items
What makes a neuron into a compass?
In 1983, James Ranck Jr. was recording from neurons in a small cortical region called the dorsal presubiculum when he noticed something strange. A cell he was monitoring would burst into activity at irregular intervals, with no obvious link to the rat's location, speed, or what the rat was looking at. Ranck watched the behavior more carefully. The pattern clicked: the neuron fired whenever the animal's head pointed in a particular compass direction — say, northeast — and went quiet the moment the head swung away. The cell didn't care where in the room the animal was. It only cared which way the animal was facing.1
Ranck reported the observation in a Society for Neuroscience abstract the following year. He called them head direction cells.2 A few years later, his postdoctoral fellow Jeffrey Taube, working with Robert Muller and Ranck at SUNY Brooklyn, published the first systematic characterization: two back-to-back papers in the Journal of Neuroscience in 1990 that defined everything from the cells' firing rates to how they responded when landmarks were rotated or removed. Those papers have since been cited over 2,600 times and remain the entry point into the field.3
Together with place cells and grid cells — which you've met in the two previous articles — head direction cells form the third pillar of the mammalian spatial navigation system. Place cells report where you are. Grid cells provide a coordinate metric. Head direction cells answer a third, logically prior question: which way am I facing?
The tuning curve: a cell with one preferred direction
The core finding is almost preternaturally clean. Each head direction cell has a single preferred firing direction — the compass heading at which it fires maximally. When a rat faces that direction, the neuron can discharge at anywhere from 5 to 115 spikes per second (mean: ~35 spikes/sec, according to the 1990 characterization). When the head rotates away, firing drops to near zero within roughly 45° of the preferred direction, and the cell is silent outside about 90° of arc. Plot firing rate on the y-axis against head direction on the x-axis, and you get a sharp triangular peak, not a broad curve.3
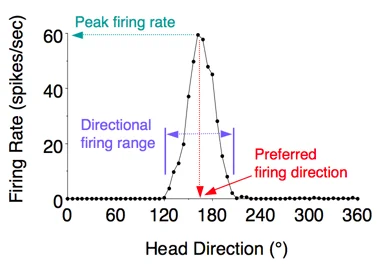
Several features make this cell type unusual. First, the preferred direction is allocentric — defined with respect to the environment, not the rat's body. A cell tuned to "northeast" keeps firing northeast whether the rat approaches from the south, sits still, or is moving fast. The cell does not encode where the head points relative to the torso; it encodes where the head points in the world. Second, among a population of head direction cells, preferred directions are distributed uniformly around the full 360°. The population holds a complete representation of all possible headings simultaneously — no direction is privileged. Third, the preferred direction is stable. Taube, Muller, and Ranck measured individual cells for days and found the same peak, at the same compass bearing, every time.
Some cells in the anterodorsal thalamus go one step further: they fire slightly in advance of the animal's actual heading, anticipating the head direction by up to 25 milliseconds.4 This "anticipatory" activity probably reflects motor efference copy signals preparing the network for an impending turn.
The circuit: from vestibule to cortex
Where does the head direction signal come from, and how does it stay accurate?
The short answer: the vestibular system, specifically the semicircular canals of the inner ear. The canals detect angular head rotation. That rotation signal travels upstream through a chain of brainstem and subcortical structures, arriving ultimately at the postsubiculum and then spreading through much of the limbic system.
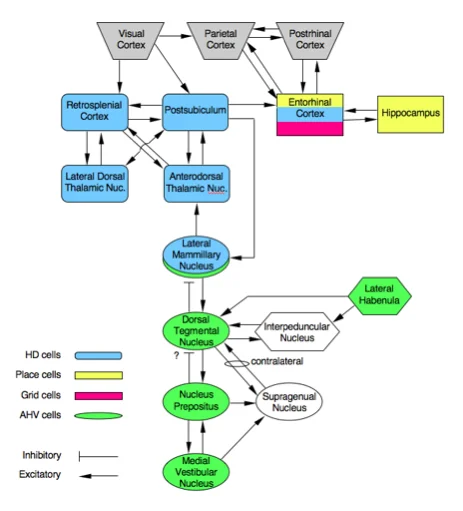
The best-understood generator node is the pair of subcortical structures: the dorsal tegmental nucleus and the lateral mammillary nuclei. Neurons in the dorsal tegmental nucleus are sensitive to angular head velocity — they fire more as rotation speed increases — and they project to the lateral mammillary nuclei, which contain the first genuine head direction cells in the ascending pathway. From the lateral mammillary nuclei, the signal goes to the anterodorsal thalamus, where head direction cells are particularly abundant and have been characterized in detail by Taube.2 From there, the signal propagates to the postsubiculum (adjacent to the hippocampus), then to the retrosplenial cortex (you met that region two articles ago), then to the entorhinal cortex, and finally back to the hippocampus, where it can be integrated with place cell firing.
Lesion experiments confirm this hierarchy. Destroying the lateral mammillary nuclei abolishes head direction cell activity in the anterodorsal thalamus. Destroying the postsubiculum does not eliminate thalamic HD cells, but it does disrupt their ability to use visual landmarks to anchor their preferred directions.2
Landmarks, darkness, and the ring attractor
That distinction — maintaining the signal versus anchoring it — is worth unpacking, because it reveals something interesting about how the system is organized.
Head direction cells can maintain their preferred directions in complete darkness. If you turn off the lights in the recording arena, the cells keep firing in the correct direction for some time. Eventually, with no external reference, the signal drifts — the "compass needle" slowly rotates away from the real-world bearing — but it doesn't simply stop. This persistence in the dark implies that the network has an internal mechanism to sustain the signal without sensory input, using vestibular and proprioceptive cues alone. The technical name for this is path integration.
The dominant theoretical framework for how this works is the ring attractor network.4 Imagine all head direction cells as points arranged around the circumference of a ring, each positioned at their preferred direction. Cells representing nearby directions excite each other; cells representing distant directions inhibit each other. The net effect is that neural activity concentrates into a single "hill" on the ring — only cells near one direction fire at any moment — and the hill can be moved around the ring by inputs from angular velocity cells. This internal architecture gives the system its momentum: once the hill settles on a direction, it stays there unless pushed.
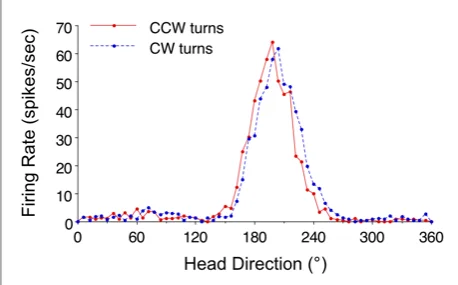
Visual landmarks, when available, act as a correction signal. When a salient cue (like the white card taped to the wall of the recording cylinder in Taube's experiments) is rotated 90°, the preferred firing directions of all head direction cells recorded simultaneously rotate by 90°. The critical observation is that all cells shift together by the same amount — their relative preferred directions stay fixed.2 The entire network behaves as a coherent ensemble, not as a collection of independent sensors. Landmarks anchor the ring; the ring attractor keeps the ensemble coherent between glimpses of landmarks.
The navigation ensemble: three cell types, one GPS
By the mid-2000s, with place cells (hippocampus, 1971), head direction cells (dorsal presubiculum, 1984), and grid cells (medial entorhinal cortex, 2005) all characterized, neuroscientists had a coherent picture of a distributed spatial navigation system.
The three cell types solve complementary problems. Place cells map the question "where am I?" Grid cells provide metric coordinates — a ruler for space. Head direction cells answer "which way am I facing?" — a prerequisite for deciding which direction to move. These three signals converge downstream: conjunctive cells in the medial entorhinal cortex fire in a grid pattern and only when the animal faces a specific direction. The hippocampus integrates all three signals to build a full allocentric map of the environment that can support both navigation and episodic memory.
Head direction cells have since been found in many species beyond rats: mice, chinchillas, monkeys, and bats. In bats — which navigate in three dimensions — the HD system operates in 3D space, not just the horizontal plane.4 Analog systems have been found in insects too: the Drosophila "ellipsoid body" contains neurons anatomically arranged along a ring that encode heading direction, and they appear to implement an attractor-like mechanism. The head direction computation may be ancient and conserved.
Whether the human brain has head direction cells is almost impossible to test with standard fMRI, which cannot resolve single neurons. But the retrosplenial cortex, which receives head direction signals from the thalamus in rodents, has been repeatedly implicated in directional orientation in human imaging studies — consistent with the idea that the system is present in our brains, even if we cannot record it directly.
Open questions
Several threads remain genuinely unresolved.
How does the system initialize? When an animal enters a completely novel environment for the first time, its head direction cells adopt some preferred direction — but which one? The initial alignment appears to be arbitrary and is then locked in when the animal associates landmarks with directions. The mechanism for this "first-pass" alignment is not fully understood.
Why does the signal degrade during inverted locomotion? Taube's lab found that head direction cells maintain coherent firing when rats climb walls — treating the vertical surface as their "floor" — but the signal breaks down when the animal is inverted, moving along the ceiling. Something about being upside down disrupts the system; otolith organ inputs (which signal gravitational direction) may be essential for resolving ambiguities that arise in non-standard orientations.
How exactly do head direction and grid cell computations interact? The lateral mammillary nucleus and anterodorsal thalamic circuit are upstream of the medial entorhinal cortex, suggesting HD signals help establish the orientation of grid cell lattices. Lesions that abolish head direction signals in the anterodorsal thalamus also disrupt grid cell regularity in the entorhinal cortex. But the precise mechanistic link — how a direction signal gets converted into the orientation angle of a hexagonal grid — remains an active research area.2
Why it matters
Head direction cells are, in a sense, the simplest of the three cell types: one neuron, one preferred heading, very little else. That simplicity made them tractable for theory. The ring attractor model built to explain them became a template for thinking about persistent neural activity more broadly — how networks sustain a representation without continuous sensory input. It has since been invoked to explain working memory, motor planning, and other forms of "internal state" that the brain must maintain across time.
For navigation specifically, head direction cells solved a conceptual puzzle. Any spatial map is useless without knowing which way you are facing. You can know you are at coordinates (4, 7) on a grid, but unless you know whether north is in front of you or behind you, you cannot use that information to plan a route. The head direction system provides the orientation reference that makes the cognitive map actionable.
Ranck's 1983 observation in the postsubiculum was one of those quietly decisive moments in systems neuroscience: a single cell, firing in a pattern that made no sense until the right variable was checked, then revealing a dedicated neural mechanism that the brain had quietly maintained across millions of years of evolution.
Landmark paper: Taube, J.S., Muller, R.U., & Ranck, J.B. Jr. (1990). Head-direction cells recorded from the postsubiculum in freely moving rats. I. Description and quantitative analysis. Journal of Neuroscience, 10(2), 420–435. 3 (PMID 2303851)
Course connection: MIT 9.13 The Human Brain, Topic 6.4 — "Head Direction Cells, Grid Cells, and Border Cells" 5
Add more perspectives or context around this Post.